Posted by Kasra
Studying mechanisms of parasitism teaches us a lot about the host physiology as well as parasite pathogenicity. Millions of years of co-evolution have adapted the parasites to exploit various homeostatic and pathologic processes of the host for their benefit. A nice example is recent study on the protozoan parasite Trypanosoma cruzi and its exploitation of host microvesicles. T. cruzi, is the causative agent of Chagas disease in Southern America’s and similar to other parasites has a complicated life-cycle. Once injected into the mammalian host, it toggles between an intracellular and bloodstream stage. In the blood-stream, among all, the parasite should fight complement-mediated lysis and inside the host cells it should avoid being killed by activated immune cells. Cestari et al. show how the parasite is able to get help from host-released microvesicles for survival in both of these stages.
Microvesicles or plasma membrane vesicles or (PMVs) are relatively large (between 200-500nm, compared to exosomes that are between 40-100nm) exovesicles released by eukaryotic cells. Not much is known about their part in homeostasis, but they have been shown to play roles in cancer and infections. More research on their role in disease can help us tell what they do in normal conditions.
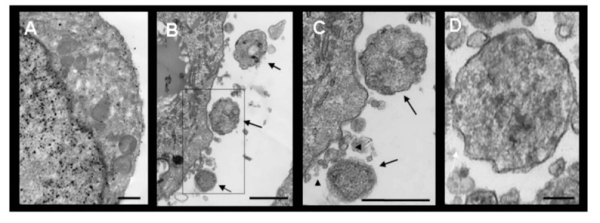
Microvesicles released from THP-1 cells, Cestari et al. 2012, J Immunology
Cestari et al. show that contact between T. cruzi metacyclic trypomastigotes and THP-1 induces a rapid augmentation in release of microvesicles from THP-1 cells in an apoptosis independent manner. They suggest that the cytoplasmic trigger for release of microvesicles is a transient increase in Ca2+ levels. Interestingly, they show that these vesicles are able to save the parasite from complement mediated lysis by binding C3 convertase on the parasite surface and inhibiting C3 cleavage.
Although they do not look at the complete content of the microvesicles, Cestari et al. show that microvesicles contain TGF-β, a generally antiproliferative and antiactivatory cytokine. Levels of TGF- β are not surprisingly different in microvesicles from different cell origins. However, once present together with the parasite, they allow a stronger infection of Vero cells (an epithelial cell line). They show that pre-incubation with microvesicles allows for a stronger T. cruzi infection and this effect is specifically due to TGF- β. This can be important for parasitism, since they have also suggested that mice infected with T. cruzi have higher levels of microvesicles in their bloodstream compared to normal mice.
Like any other exciting piece of research, this study raises a lot of questions. For instance, why is there an increase in microvesicle release following contact with T. cruzi? Is this a common effect with other pathogens or eukaryotic parasites? What is the role of these vesicles when they are not being exploited? What other cytokines/signaling proteins do they contain? What do they do? What are their target cells? And so on. I am very eager to read the follow-up papers and learn more about this method of cell-cell communication.
Cestari I, Ansa-Addo E, Deolindo P, Inal JM, & Ramirez MI (2012). Trypanosoma cruzi Immune Evasion Mediated by Host Cell-Derived Microvesicles. Journal of immunology (Baltimore, Md. : 1950), 188 (4), 1942-52 PMID: 22262654
![]()
